
Spurensuche im Stammbaum
In jeder Ahnentafel gibt es besondere Punkte, die es wert sind, genauer betrachtet zu werden. Unter einer historischen Perspektive sind das das erste Auftreten einer Verwandtenehe, die ersten „Ahnengeschwister“, der erste primäre Ahnenimplex, die Person mit der höchsten Ahnennummer usw. Unter einer phylogenetischen Perspektive begegnen uns in jeder Ahnentafel LUCA, LECA, der „Adam des Y-Chromosoms“, die „mitochondriale Eva“ oder der „Most Recent Common Ancestor“ (MRCA). Was es damit auf sich hat, erklärt unsere neue Info-Grafik.
Von Heiko Hungerige
Wer sich mit Familienforschung beschäftigt, stößt früher oder später auf besondere Stellen in der eigenen Ahnentafel, die neugierig machen: das erste Auftreten einer Verwandtenehe, die Person mit der höchsten Ahnennummer oder die ersten „Ahnengeschwister“, die dieselben Vorfahren teilen (primärer Ahnenimplex). Doch auch aus genetischer Sicht gibt es spannende (wenn auch mit den Methoden der Geschichtswissenschaft nicht mehr erforschbare) „Fixpunkte“ wie die „mitochondriale Eva“ oder den „Adam des Y-Chromosoms“. In diesem Beitrag werfen wir einen genaueren Blick auf diese Schlüsselpositionen in einer Ahnentafel – dort, wo sich Geschichte und Genetik berühren.
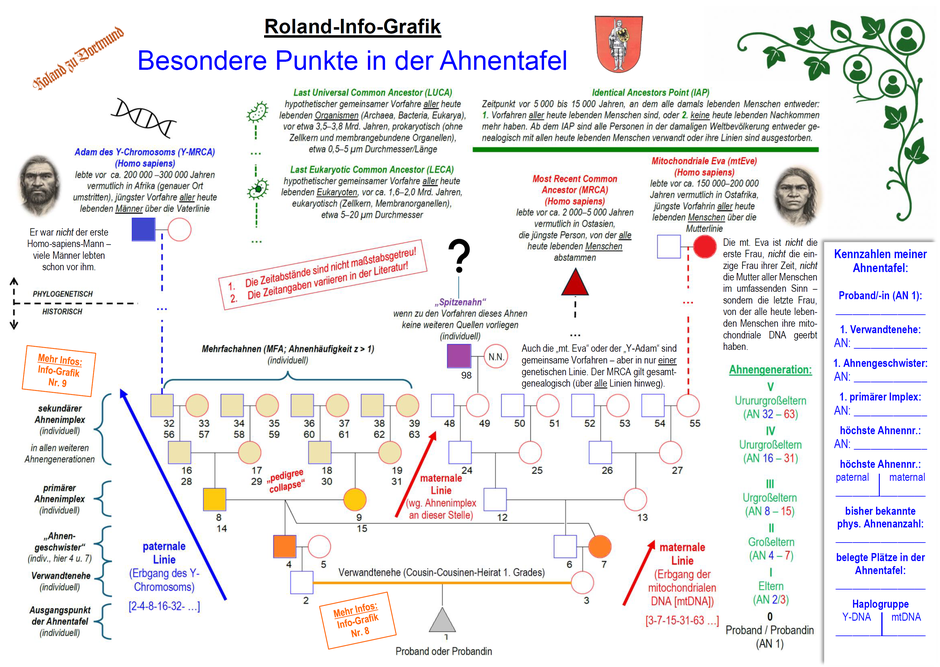
Unsere neue Roland-Info-Grafik: Besondere Punkte in der Ahnentafel. (Für den Download der Info-Grafik auf das Bild klicken.)
Die historische Perspektive
Ahnenforschung ist immer „monozentrisch“, wie Siegfried Rösch 1956 in seinem Buch „Goethes Verwandtschaft“ bemerkte: Ausgangspunkt einer jeden Ahnentafel ist ein Proband oder eine Probandin mit
der Ahnennummer (AN) 1; sie sind der „Nullpunkt“ dieser „stets ins Unendliche entgleitenden[n], unfaßbar scheinende[n] Masse“ an Vorfahren (Rösch, 1956, S. 3; vgl. Info-Grafik Nr. 1).
Bei einer Cousin-Cousinen-Ehe 1. Grades in der Elterngeneration treten die ersten „Ahnengeschwister“ bereits in der Großelterngeneration des/der Prob. auf: In unserer Info-Grafik
ist der Großvater väterlicherseits (AN 4) ein Bruder der Großmutter mütterlicherseits (AN 7). Oder anders gesagt: Die Braut (AN 3) hat den Sohn (AN 2) des Bruders (AN 4) ihrer Mutter (AN 7)
geheiratet (eine sog. Kreuzcousinenheirat, lat.: amitini; vgl. Info-Grafik Nr. 8).
Die Eltern dieser „Ahnengeschwister“ treten dann in der Urgroßelterngeneration des/der Prob. mit den Ahnennummern 8 und 14 sowie 9 und 15 notwendigerweise doppelt auf
(Mehrfachahnen), besetzen also jeweils zwei Plätze in der Ahnentafel. Ein primärer Ahnenimplex (= Ahnengleichheit) ist entstanden, der sich in den weiteren
Ahnengenerationen als sekundärer Ahnenimplex (weil biologisch zwangsläufig) fortsetzt. Die Ahnentafel „kollabiert“ also in gewisser Weise (weswegen der Ahnenimplex im Englischen
auch pedigree collapse genannt wird; vgl. Info-Grafik Nr. 9).
Da Cousin-Cousinen-Heiraten 1. Grades nach dem Kirchenrecht verboten sind (das ist noch heute so, nach dem bürgerlichen Gesetz (BGB) sind sie dagegen erlaubt), ist der Generationsabstand zwischen
der Verwandtenehe und dem ersten Auftreten von „Ahnengeschwistern“ als Auslöser für den Ahnenimplex in der Regel sehr viel größer; meistens liegen so viele Generationen dazwischen, dass die
Brautleute oft gar nicht wussten, dass sie (mehr oder weniger entfernt) miteinander „verwandt“ sind.
Verwandtenehen, Ahnengeschwister und Ahnenimplex kommen ausnahmslos in jeder Ahnentafel vor; wer sie noch nicht gefunden hat, hat nicht lange genug danach gesucht. Dies ist
notwendigerweise so, weil die physikalische (tatsächliche) Anzahl der Ahnen immer deutlich kleiner ist als die theoretische Anzahl (die sich ja mit jeder Ahnengeneration verdoppelt).
Jede individuelle Ahnentafel lässt sich also durch diese „besonderen Ahnen“ charakterisieren, zu denen auch die sog. „Spitzenahnen“ zählen, also Personen, zu deren Vorfahren keine weiteren Quellen vorliegen („tote Punkte“, in unserem Beispiel: AN 98).
Wer will, kann in der rechten Spalte unserer Info-Grafik diese Kennzahlen für die eigene Ahnentafel notieren:
- Den/die Probanden/-in als Ausgangspunkt der Ahnentafel (Namen ausschreiben).
- Das Auftreten der ersten Verwandtenehe (in unserem Beispiel: AN 2 u. 3).
- Das Auftreten der ersten „Ahnengeschwister“ als Auslöser für den Implex (in unserem Beispiel: AN 4 u. 7).
- Das Auftreten des ersten primären Ahnenimplexes (in unserem Beispiel die Mehrfachahnen 8/14 und 9/15).
- Die höchste Ahnennummer in der gesamten Ahnentafel (soweit bisher bekannt; in unserem Beispiel: AN 63).
- Die höchste Ahnennummer in der paternalen (Erbgang des Y-Chromosoms) und maternalen (Erbgang der mitochondrialen DNA) Ahnenlinie (in unserem Beispiel: AN 32 (= 56) und 63 (= 39).
- Die bisher bekannte physikalische (tatsächliche) Ahnenanzahl (Mehrfachahnen werden nur einmal gezählt).
- Die belegten Plätze in der Ahnentafel (Mehrfachahnen zählen gemäß ihrem Vorkommen).
- Und wer DNA-Genealogie betreibt, kann zusätzlich die Haplogruppen für das Y-Chromosom (Y-DNA) und für die mitochondriale DNA (mtDNA) notieren.
Die phylogenetische Perspektive
Doch selbst, wenn sich einige Ahnenlinien bis in die karolingische Zeit zurückverfolgen lassen: Irgendwann versiegen die historischen Quellen (oder werden unglaubwürdig). Die Ahnenlinien gehen aber selbstverständlich weiter: Die Abstammungslinie des Homo sapiens verläuft über viele Vorformen, beginnend bei frühen Primaten (vor ca. 60 Mio. Jahren) über verschiedene Homininen-Arten bis hin zum modernen Menschen (seit ca. 300.000 Jahren) – ein Prozess, geprägt von Anpassung, Migration und genetischer Vielfalt.

Vor etwa 50.000 bis 100.000 Jahren lebten mindestens fünf bis sieben verschiedene Menschenarten gleichzeitig auf der Erde. Das macht deutlich, dass Homo sapiens ursprünglich nur eine Menschenart unter mehreren war – mit teilweise sogar genetischem Austausch, wie DNA-Analysen zeigen (z. B. Neandertaler-Gene in heutigen Menschen). Die Vorstellung vom „einsamen modernen Menschen“ ist also historisch nicht haltbar. (Bilder: Wikimedia Commons)

Angaben von Svante Pääbo, Direktor am Max-Planck-Institut für evolutionäre Anthropologie in Leipzig (in der Literatur teilw. abweichende Angaben)
Auch in dieser, hypothetisch „erweiterten Ahnentafel“ treffen wir auf markante Punkte. Als erster begegnet uns auf unserer Zeitreise in die Vergangenheit der Most Recent Common Ancestor
(MRCA), ein Homo sapiens, der vor 2.000 bis 5.000 Jahren vermutlich in Ostasien lebte. Er ist die jüngste Person, von der alle heute lebenden Menschen abstammen und damit die zuletzt
lebende Person, die in jedem heutigen Familienstammbaum vorkommt – bei Ihnen, bei mir, bei allen. Bemerkenswert ist, dass er nicht in der fernen Urgeschichte, sondern vermutlich vor wenigen
Tausend Jahren lebte. Er ist damit ein faszinierendes Beispiel dafür, wie eng die Menschheit genealogisch miteinander verwoben ist.
Interessant in diesem Zusammenhang ist auch der sog. Identical Ancestors Point (IAP). Das ist der Zeitpunkt vor 5.000 bis 15.000 Jahren, an dem alle damals lebenden Menschen
entweder:
- Vorfahren aller heute lebenden Menschen sind, oder
- keine heute lebenden Nachkommen mehr haben.
Ab dem IAP sind alle Personen in der damaligen Weltbevölkerung entweder genealogisch mit allen heute lebenden Menschen verwandt oder ihre Linien sind ausgestorben. Der Oxforder Bioinformatiker
Jotun Hein (2004) formuliert es noch drastischer: Angenommen, Sie kommen um 3000 v. Chr. in ein Dorf, dann ist der erste beliebige Mensch, dem Sie begegnen, wahrscheinlich ein Vorfahre von
Ihnen!
Gehen wir noch weiter in der Zeit zurück, begegnen uns die „mitochondriale Eva“ (mtEve) und der „Adam des Y-Chromosoms“ (Y-Adam oder Y-MRCA). Auch sie sind gemeinsame Vorfahren, aber anders als
der MRCA nur in je einer einzigen genetischen Linie, nämlich der paternalen (Y-Adam) bzw. maternalen (mtEve) Ahnenlinie. Sie sind symbolisch bedeutsam, aber genealogisch nur
Teilausschnitte eines viel größeren Stammbaums.
Die mitochondriale Eva (mtEve) ist die jüngste Frau, von der alle heute lebenden Menschen über eine ausschließlich mütterliche Linie (Mutter → Tochter → …)
abstammen. Nur Frauen können diese Linie biologisch weitergeben (da nur Eizellen Mitochondrien vererben). Sie ist nicht die alleinige Vorfahrin aller Menschen, aber Teil des Stammbaums aller und
lebte vor ca. 150.000 bis 200.000 Jahren vermutlich in Ostafrika (die Jahresangaben werden in der Fachliteratur unterschiedlich geschätzt). Die häufigsten mitochondrialen Haplogruppen in
Westeuropa sind H (ca. 40–45 %), U (ca. 10–15 %), J (ca. 5–10 %), T (ca. 5–10 %) und K (ca. 5–8 %).
Der Adam des Y-Chromosoms (Y-MRCA) ist der jüngste Mann, von dem alle heute lebenden Männer über eine ausschließlich väterliche Linie (Vater → Sohn → …)
abstammen. Frauen erben kein Y-Chromosom, sind also nicht seine Nachkommen in dieser Linie. Sie können aber indirekt von ihm abstammen, z. B. über ihre Väter, Großväter etc., allerdings nicht
über eine rein männliche Linie. Der Y-Adam lebte vor ca. 200.000 bis 300.000 Jahren vermutlich in Afrika (genauer Ort umstritten). Auch er war nicht der erste Homo-sapiens-Mann – viele Männer
lebten schon vor ihm. Der „Adam des Y-Chromosoms“ wird heute mit der Haplogruppe A0-T (oder älter: A00) in Verbindung gebracht. Alle anderen männlichen Linien sind aus dieser
Ursprungs-Haplogruppe hervorgegangen. Die in Westeuropa am häufigsten vorkommende Y-Haplogruppe ist R1b (R-M343), die vermutlich vor 4.500 bis 5.000 Jahren aus dem heutigen Südrussland und der
Ukraine mit den indoeuropäischen Yamna-Kulturen nach Westeuropa kam.
Diese beiden „Ahnen“ lebten nicht zur gleichen Zeit und stehen für unterschiedliche Abstammungslinien in der menschlichen Evolutionsgeschichte. Zu beachten ist: Weder die mitochondriale Eva noch
der Y-MRCA sind die einzigen gemeinsamen Vorfahren aller Menschen oder die ältesten ihrer Art oder die einzigen lebenden Menschen zu ihrer Zeit. Jeder heutige Mensch
hat tausende andere Vorfahren (nämlich „die Mitte“ einer Ahnentafel), die nicht in direkter mtDNA- oder Y-Linie liegen, aber ebenso entscheidend sind.
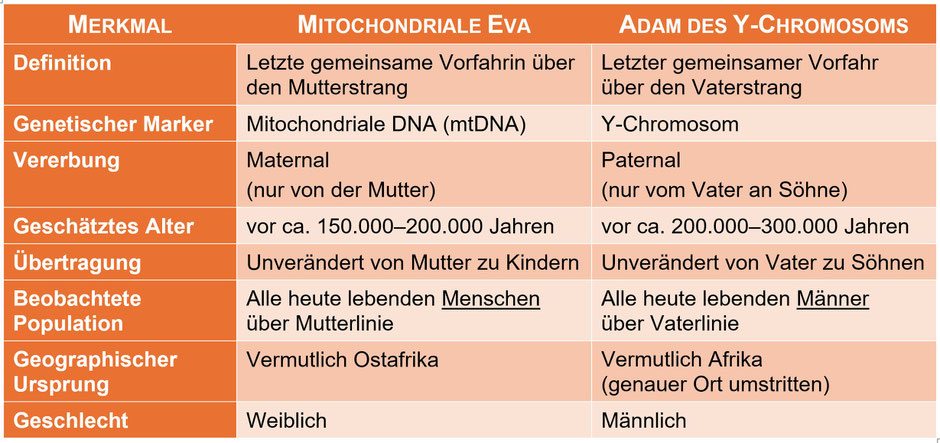
Vergleich mitochondriale Eva vs. Adam des Y-Chromosoms
Gehen wir noch viel weiter zurück, nämlich in die Zeit vor ca. 1,6–2,0 Milliarden Jahren, treffen wir auf LECA, den Last Eukaryotic Common Ancestor, also den
gemeinsamen Vorfahren aller heute lebenden Eukaryoten, Lebewesen mit echtem Zellkern.
LECA war eine vollwertige, komplexe eukaryotische Zelle mit etwa 5–20 µm Durchmesser – mit Kern, Membranorganellen (inkl. Proto‑Mitochondrien), einem dynamischen Cytoskelett und den Grundzügen
von Phagozytose und Zellteilung. Seine Ausstattung bildete bereits alle zentralen Merkmale ab, die wir heute in Pilzen, Pflanzen, Tieren und Protisten (alle ein- bis wenigzelligen Eukaryoten,
also Protozoen, Mikropilze und (eukaryotische) Mikroalgen) wiederfinden. LECA lebte in einer mikrobiell dominierten, aquatischen Welt mit steigendem Sauerstoffgehalt, inmitten vielfältiger
Einzeller. Seine Umgebung begünstigte Symbiosen, Energiegewinnung, Kompartimentierung – entscheidende Schritte auf dem Weg zu allen heutigen Eukaryoten, also zu Pflanzen, Tieren und Menschen.
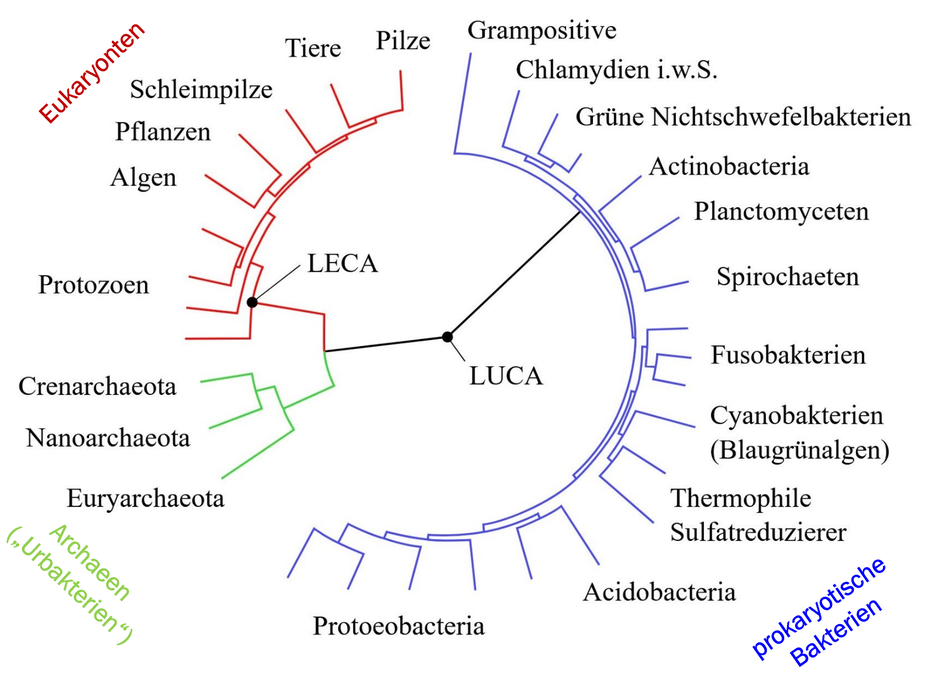
Moderne Darstellung der Verwandtschaftsverhältnisse aller rezenter Organismen in Form eines sogenannten Kladogramms (in diesem Umfang auch „Tree of Life“, „Baum des Lebens“ genannt). Der Urvorfahr (LUCA) steht an der Wurzel dieses Kladogramms. (Quelle: Von TimVickers (‘original’ PNG)User_A1 (SVG conversion of ‘original’ PNG)Gretarsson (derivative work based on the SVG)also see file description of the source SVG - CollapsedtreeLabels-simplified.svg, CC0, https://commons.wikimedia.org/w/index.php?curid=70882841)
Doch damit sind wir noch nicht am Ende unserer Reise durch die Zeit: Vor etwa 3,5–3,8 Mrd. Jahren, also noch weit vor LECA, lebte LUCA, der Last Universal Common
Ancestor. Er ist der hypothetische gemeinsame Vorfahre aller heute lebenden Organismen (Archaea, Bacteria, Eukarya).
Er war klein und einfach, aber nicht primitiv. Mit vermutlich 0,5–1 Mikrometer Durchmesser besaß er keinen Zellkern, aber RNA und DNA als Erbinformation und verfügte auch über Ribosomen, war also
fähig zur Proteinbiosynthese. Der Lebensraum von LUCA war vermutlich sauerstofffrei oder sauerstoffarm, z. B. heiße Tiefseequellen. Sauerstoff war in der frühen Erde noch kaum vorhanden – die
sogenannte „Große Sauerstoffkatastrophe“ (Entstehung freier O₂ durch Cyanobakterien) kam erst ca. eine Milliarde Jahre später. LUCA war daher mit sehr hoher Wahrscheinlichkeit anaerob, verfügte
aber bereits über erste Enzyme, die mit Sauerstoff umgehen konnten, auch wenn dieser noch selten war.
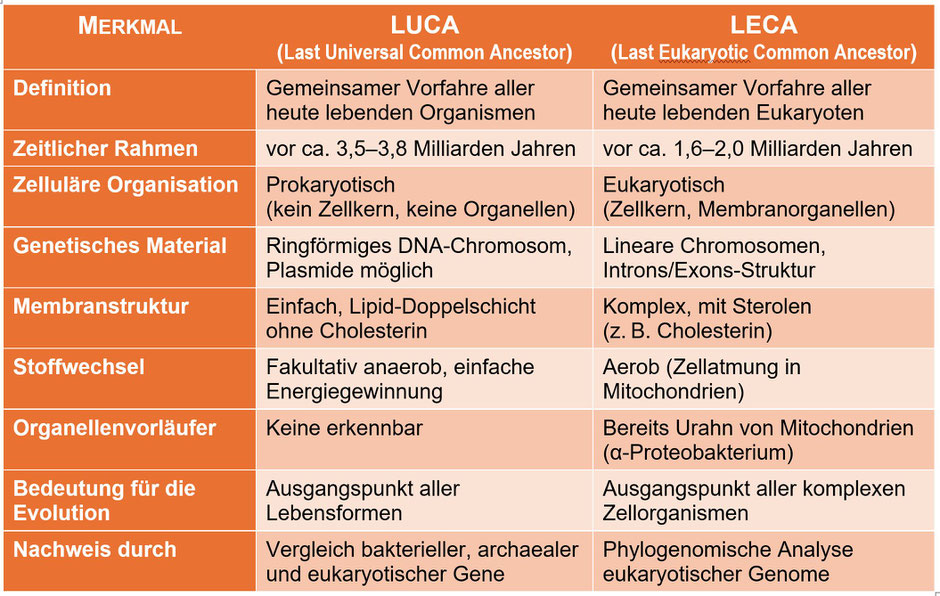
Vergleich LUCA vs. LECA. Zur Erklärung: Plasmide sind in der Regel ringförmige, autonom replizierende, doppelsträngige DNA-Moleküle. Exons sind die codierenden Abschnitte eines Gens – sie enthalten die Anleitung zur Herstellung von Proteinen. Introns sind die nicht-codierenden Abschnitte – sie werden zunächst mitgelesen, aber beim Spleißen wieder entfernt, bevor das Protein entsteht. Der Übergang zur Intron-/Exon-Struktur ist ein Schlüsselmerkmal der Eukaryoten. „Fakultativ anaerob“ bedeutet, dass LUCA ohne Sauerstoff leben konnte, aber vermutlich in der Lage war, Sauerstoff zu nutzen, wenn er vorhanden war. Eine phylogenomische Analyse ist eine Methode der Bioinformatik und Evolutionsbiologie, bei der man mithilfe genomweiter Daten (also sehr vieler Gene oder ganzer Genome) evolutionäre Verwandtschaften zwischen Organismen untersucht.
Besondere Punkte in der Ahnentafel helfen, komplexe Strukturen familiärer Herkunft besser zu verstehen. Sie erklären etwa das Phänomen der Mehrfachahnen, zeigen, wie eng alle Menschen
genealogisch miteinander verwoben sind, und markieren die Grenzen historisch belegbarer Linien. Gleichzeitig erweitern sie den Blick auf die Ursprünge des Lebens und verdeutlichen, dass
Genealogie nicht nur persönliche Familiengeschichte, sondern auch Teil der Menschheitsgeschichte ist.
P.S.: Wer mehr „Geschichten vom Ursprung des Lebens“ lesen will, sei auf das gleichnamige Buch von Richard Dawkins aus dem Jahr 2008 verwiesen (bei Ullstein): Ausgehend von der
landwirtschaftlichen Revolution am Ende der letzten Eiszeit vor etwa 10.000 Jahren bis hin zu den Archaea und Eubakterien der fernen Vergangenheit begibt sich der Autor auf eine faszinierende
„Zeitreise auf Darwins Spuren“. Sein neuestes Werk, „Das große Buch der Evolution“, stellt der wohl bekannteste noch lebende Evolutionsbiologe am Freitag, den 7.11.2025 um 20 Uhr im Kleinen Saal
der Laeiszhalle in Hamburg vor („Ein Abend mit Richard Dawkins“).
Alle Roland-Info-Grafiken können hier heruntergeladen werden: