
Analysiert mit der genealogischen Software "AhnenImplex" – Ein Beispiel
In diesem Blog-Beitrag werden an einem Beispiel einige der Analysemöglichkeiten der genealogischen Software AhnenImplex vorgestellt. Der Blog-Beitrag kann als Ergänzung zu dem in diesem Monat in Heft 3 (S. 18-20) der „Computergenealogie“ erschienenen Artikel „Quantitative Genealogie: Verwandtschaft messbar machen mit AhnenImplex“ gelesen werden.
Ausgangspunkt der Analyse ist eine umfangreiche GEDCOM-Datei von Gerhard Kreile mit über 18.000 Personen. Von ihm sind derzeit (2023) 529 Vorfahren bekannt, darunter neun Vollgeschwister-
und zwei Halbgeschwister-Paare („Ahnengeschwister“). Deren Eltern (bzw. Elternteile bei den Halbgeschwistern) tauchen in der Ahnentafel mehrfach auf (primärer Ahnenimplex).
Die genealogische Software AhnenImplex wurde von Martin Jülich (Chemnitz) auf Anregung von Arndt Richter (München) speziell dafür entwickelt, Mehrfachahnen (MFA) in Ahnentafeln
(AT) zu finden und hinsichtlich statistisch-genealogischer Kennwerte wie Ahnenhäufigkeit (z), Ahnenimplex (ik), biologischem Verwandtschaftsgrad (b bzw. g’b), Verwandtschaftskoeffizient (R),
Generationsverschiebung (gb) u.v.m. zu analysieren. Grundlage hierfür ist die Quantitative Genealogie von Siegfried Rösch (1899 – 1984).
Von Gerhard Kreile und Heiko Hungerige
Mehrfachahnen und Ahnenimplex - was war das nochmal?
In jeder ausreichend erforschten Ahnentafel tritt früher oder später aus biologischer Notwendigkeit Ahnenimplex oder „Ahnengleichheit“ auf, früher – etwas missverständlich – auch als „Ahnenschwund“ oder „Ahnenverlust“ bezeichnet. Gemeint ist damit, dass mindestens zwei oder auch mehrere Positionen in der Ahnentafel durch ein und dieselbe Person besetzt werden; diese treten in der Ahnentafel des Probanden dann doppelt oder mehrfach auf (Mehrfachahnen).
Ausgelöst wird jeder Ahnenimplex (ik) zwangsläufig durch ein Geschwisterpaar (oder auch mehrere Geschwister), deren Nachfahren in einer der folgenden Generationen heiraten
(Verwandtenehen) und schließlich (oft erst nach mehreren Generationen) den Probanden hervorbringen. Diese Ahnengeschwister nehmen „bei der Berechnung verwandtschaftlicher Verflechtungen
eine Daten- und Brückenfunktion ein“ (Richter, 1997, S. 61). Konkret bedeutet dies, dass allein aus der Zusammenstellung dieser Ahnengeschwister in einer sog. Verschwisterungsliste (VSL;
vgl. Tab. 1) verschiedene statistisch-genealogische Kennwerte abgeleitet werden können, u. a. sämtliche Ahnennummern (Kekule) der Ahnentafel, der biologische Verwandtschaftsgrad und der
Ahnenimplex.
Verschwisterungsliste (VSL)
Mit AhnenImplex lässt sich leicht eine solche Verschwisterungsliste erstellen. Tabelle 1 zeigt die den Ahnenimplex auslösenden Voll- und Halbgeschwister in einer Übersicht. Ebenfalls angegeben sind die jeweils kleinsten Ahnennummern der Ahnengeschwister sowie die jeweils „ausgefallenen Ahnennummern“ (Geschwistereltern). Was damit gemeint ist, soll nachfolgend an zwei Beispielen verdeutlicht werden.
In der väterlichen Ahnenlinie Kreile lösen die Vollgeschwister Valentin (* 1620) und Adam (* 1612) aus Geroda (Teil des Marktes Geroda im Landkreis Bad Kissingen, Unterfranken) den Ahnenimplex aus; beide Geschwister sind Vorfahren des Probanden. Sie treten in seiner AT mit den AN 512 und 1136 auf. Die Eltern dieser Ahnengeschwister, Adam Kreile († 1637) und Catharina Fell (ca. 1580 – 1637), tauchen in der AT erstmals mit den AN 1024 und 1025 als Eltern von Sohn Valentin (AN 512) auf, dann aber nochmal mit den AN 2272 und 2273 als Eltern von Sohn Adam (AN 1136). Die Personen mit den AN 2272 und 2273 sind also in dem Sinne „ausgefallen“, als sie schon mit den AN 1024 und 1025 in der Ahnentafel vorhanden und somit „erforscht“ sind. Nicht ausgefallen sind sie aber in biologischer Hinsicht, im Gegenteil: Da sie doppelt in der AT auftauchen (wegen einer weiteren Verwandtenehe in der Nachkommenschaft tatsächlich sogar dreifach), geben sie auch ihre Gene zwei Mal (bzw. drei Mal) an die nachfolgenden Generationen weiter. Die Wahrscheinlichkeit, dass sich Teile ihres Genoms in den nachfolgenden Generationen erhalten, verdoppelt (bzw. verdreifacht) sich also.
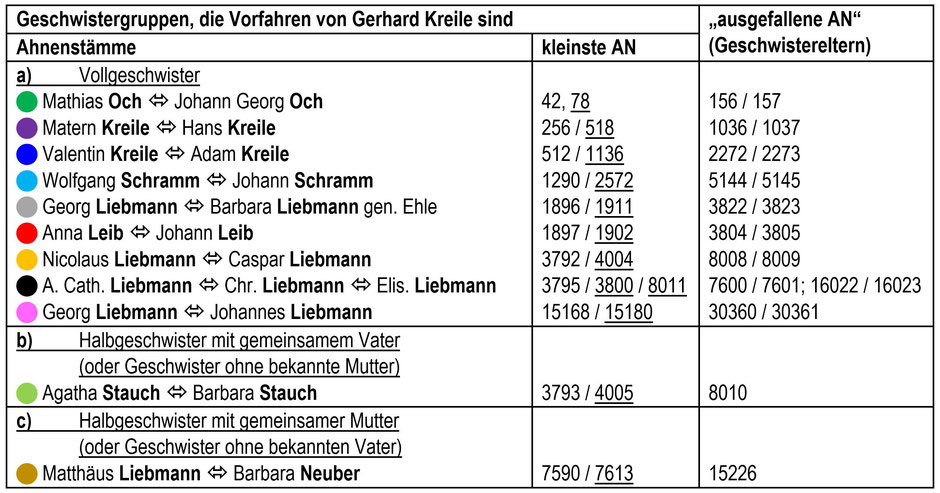
Tab. 1: Verschwisterungsliste (VSL) mit den „ausgefallenen AN“ (Implex) der Geschwistereltern in der Ahnentafel von Gerhard Kreile
Etwas anders sieht es aus, wenn der Ahnenimplex nicht durch Voll-, sondern durch Halbgeschwister ausgelöst wird: In der mütterlichen Ahnenlinie Stauch (Tab. 1, b) ist das bei den Halbgeschwistern Agatha (1605 – 1682; AN 3793) und Barbara (1627 – 1680; AN 4005) der Fall. Ihr Vater Hans Stauch (1577 – 1640) war zwei Mal verheiratet; Agatha und Barbara (beide wiederum Vorfahrinnen des Probanden) haben deswegen verschiedene Mütter. Hans Stauch tritt daher als Vater der Halbgeschwister mit den AN 7586 und 8010 in der Ahnentafel auf; im Sinne der „Erforschtheit“ ist die AN 8010 daher „ausgefallen“. Da die Halbgeschwister aber verschiedene Mütter haben (also unterschiedliche Personen), fällt von diesen keine AN aus.
Ahnenimplex (ik)
Wird einmal ein Ahnenimplex durch zwei oder auch mehrere Ahnengeschwister ausgelöst, setzt er sich in den aufsteigenden Ahnengenerationen zwangsläufig fort; auch dort treten dann also Mehrfachahnen auf. Da dies eine logische Notwendigkeit und in diesem Sinne „nichts Besonderes“ ist, wurde bereits 1907 (Artikel hier) von dem Baseler Journalisten und Genealogen Julius Oscar Hager (1853 – 1914) zwischen dem auslösenden Original-Implex und dem sich in höheren Generationen „mechanisch“ fortsetzenden Konsekutiv-Implexen unterschieden. Griffiger erscheint hier jedoch die von Arndt Richter eingeführte Unterscheidung primärer vs. sekundärer Implex. Bei der Berechnung des Ahnenimplex‘ ist daher darauf zu achten, dass nur die auslösenden primären Implexe in die Berechnung eingehen. Berücksichtigt man auch die zwangsläufig auftretenden Sekundärimplexe, wird die Angabe für den Ahnenimplex massiv überschätzt. Abbildung 1 zeigt die Zunahme des Ahnenimplex‘ (ik) in Prozent über 20. Generationen (dem Probanden Gerhard Kreile wird die Ahnengeneration „0“ zugewiesen).
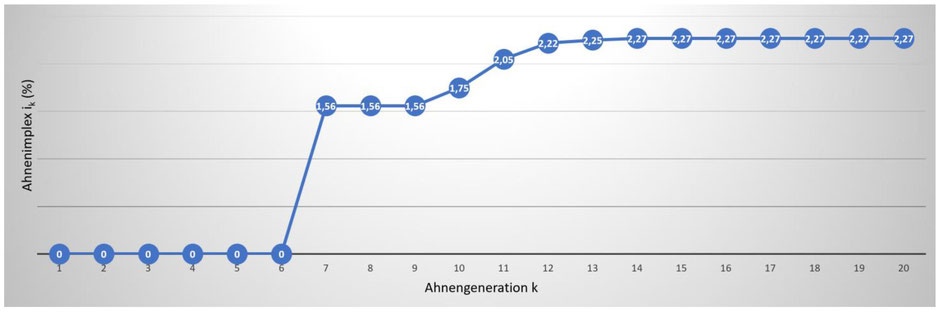
Abb. 1: Zunahme des Ahnenimplex ik über k = 20 Generationen in der Ahnentafel von Gerhard Kreile. Der Ahnenimplex erhöht sich nur in den Generationen, in denen tatsächlich auch Ahnen „ausfallen“. In den Generationen k = 8 und k = 9 sowie k = 15 bis k = 20 bleibt der Implex im Vergleich zu der jeweils vorhergehenden Generation gleich, da nur sekundärer Implex auftritt.
Obwohl auch in der 15. bis 20. Ahnengeneration Mehrfachahnen auftauchen, steigt der primäre Ahnenimplex ab der 14. Ahnengeneration nicht weiter an und erreicht in dieser mit 2,27% sein Maximum.
Insgesamt tritt (wie bereits aus der VSL zu sehen; vgl. Tab. 1) elf Mal Ahnenimplex auf, ausgelöst durch acht Vollgeschwister-Paare, ein Vollgeschwister-Triplet (Liebmann) und zwei Halbgeschwister-Paare. Veranschaulichen lässt sich dies durch einen Auszug der „geschlossenen Heiratskreise“ aus der Ahnentafel (Abb. 2), die insgesamt 529 Vorfahren des Probanden umfasst.

Abb. 2: Geschlossene Heiratskreise in der Ahnentafel von Gerhard Kreile. Angegeben sind die Ahnennummern nach Kekule. Ahnengeschwister sind farbig markiert.
Zur besseren Veranschaulichung lässt sich dieses „Ahnennetz“ in einzelne, sog. geschlossene Heiratskreise „zerlegen“. Für einen oder mehrere solcher „geschlossenen Heiratskreise“ wurde 1907 von Julius Oscar Hager (1853 – 1914) der Begriff Deszentorium eingeführt. Gemeint ist damit ein Auszug aus der AT, der die Abstammung des Probanden „von einem Ahnen in allen Linien dartut“ (Forst-Battaglia, 1913, S. 11). Die nachfolgenden Abbildungen zeigen solche Deszentorien in der AT des Probanden für die Nachfahrenlinien Kreile, Schramm und Reinhardt.

Abb. 3: Deszentorien
Mehrfachahnen (MFA)
Insgesamt treten in der AT des Probanden neun namentlich bekannte Mehrfachahnen (MFA) mit einer Ahnenhäufigkeit von jeweils z = 7, zwei MFA mit z = 5, vier MFA mit z = 4, 14 MFA mit z = 3 und 24 MFA mit z = 2 auf.
Der erste MFA mit einer Ahnenhäufigkeit von z = 7 ist Hans Liebmann (ca. 1495 – 1570) aus Geiersthal (Ortschaft im Ortsteil Lichte der Stadt Neuhaus am Rennweg im Landkreis Sonneberg,
Thüringen) mit den AN 30336, 30360, 30400, 30440, 30576, 32032 und 64088. Seine Linie lässt sich bis zu seinem Urururgroßvater Kirsten Oberländer (* ca. 1370) zurückverfolgen;
alle Vorfahren von Hans Liebmann in dieser Linie haben ebenfalls eine Ahnenhäufigkeit von z = 7 (sekundärer Implex). Kirsten Oberländer ist auch der Ahn
mit der höchsten Ahnennummer (970776) in der 20. Ahnengeneration.
Ehen naher Verwandter (konsanguine Ehen)
Neben dem Ahnenimplex ik (auch Ahnenverlustkoeffizient (AVK) genannt) wird bei Verwandtschaftstafeln auch gerne der Inzuchtkoeffizient F (coefficient of inbreeding) angegeben. Auch dieses, erstmals 1922 von dem Populationsgenetiker Sewall G. Wright (1889 – 1988) vorgestellte Maß kann ausschließlich aus der Kenntnis der oben beschriebenen Verschwisterungslisten (VSL) berechnet werden.
Autosomale Gene liegen immer paarweise vor (Allele). Ein Allel erhält das Kind von seinem Vater, das andere von seiner Mutter. Allele sind also unterschiedliche Varianten eines Gens, die sich am gleichen Genort (Locus) der jeweiligen Chromosomenpaare befinden. Erhält ein Kind identische Allele von Vater und Mutter, ist es bezüglich dieses Gens homozygot, erhält es unterschiedliche Allele ist es bezüglich dieses Gens heterozygot. Der Inzuchtkoeffizient F beschreibt nun die „Wahrscheinlichkeit, dass eine homozygote Person beide Allele eines Gens von einem einzigen gemeinsamen Vorfahren der Eltern geerbt hat“ (Murken et al., 2017, S. 255). Er ist also ein Wahrscheinlichkeitsmaß dafür, „dass sich bei Nachkommen von bereits eng biologisch verwandten Eltern dieselbe (zufällig ausgewählte) Erbinformation findet wie bei dem letzten gemeinsamen Vorfahren der beiden Elternteile.“ (Wikipedia)

Der Inzuchtkoeffizient F entspricht durchschnittlich der Hälfte des Verwandtschaftskoeffizienten R (coefficient of relationship) der beiden Elternteile zueinander, da sie jeweils nur 50 % ihrer Gene vererben (R = 2F). Er gibt also die Nähe der biologischen Verwandtschaft zweier Lebewesen anhand der Wahrscheinlichkeit an, dass sie dieselbe (zufällig ausgewählte) Erbinformation (dieselbe Kopie eines Allels) voneinander oder einem gemeinsamen Vorfahren geerbt haben. Zwischen einem Elternteil und seinem leiblichen Kind sowie zwischen Vollgeschwistern besteht ein Verwandtschaftskoeffizient von R = 0,5 (½), zwischen Halbgeschwistern sowie Großeltern und Enkel nur noch ein Verwandtschaftskoeffizient von R = 0,25 (¼).
Ein Beispiel: In den Nachfahrenlinien Conrad Schramm (* ca. 1570, AN 2580 und 5144, z = 2, väterliche Seite) liegt die Heirat der miteinander verwandten Ehepartner Johann Georg Hülß (1697 – 1776; AN 160) und Eva Catharina Schleicher (1705 – 1775; AN 161) fünf bzw. vier Generationen nach dem Implex (ausgelöst durch die „Ahnenbrüder“ Johann und Wolfgang Schramm, zwei Söhnen von Conrad Schramm).
Die Eheleute waren also Ururur- bzw. Ururenkelkinder (Generationsverschiebung) ihres gemeinsamen Vorfahren Conrad Schramm und sich ihrer „Verwandtschaft“ wohl noch nicht einmal bewusst. Ihr gemeinsamer Sohn Johann Nicolaus Hülß (1738 – 1802; AN 80) war also ein Kind aus einer Verwandtenehe und hat demnach einen Inzuchtkoeffizienten von F = 0,0019 sowie einen Verwandtschaftskoeffizienten von R = 0,0038.
Er hat also eine Wahrscheinlichkeit von 0,19% für irgendein Allel seines Vorfahren Conrad Schramm homozygot zu sein (Abb. 4). Da das Basisrisiko für ein Kind mit einer nicht vorhersagbaren genetischen Erkrankung für die Allgemeinbevölkerung mit 2-4% angegeben wird, gilt dies als genetisch unbedenklich. Bei einer Cousin-Cousinen-Heirat 1. Grades erhöht sich dieses Risiko für das Kind aber beispielsweise auf das Doppelte (6,25%) und liegt damit über dem Basisrisiko.
Abb. 4: Inzuchtkoeffizient von Johann Nicolaus Hülß
Generationsverschiebung und Generationsspektren (gb)
Durch unterschiedliche Generationsabstände in den einzelnen Nachfahrenlinien kommt es häufiger vor, dass ein Mehrfachahn in verschiedenen Generationen in Erscheinung tritt (Generationsverschiebung). Für die Notation verwendet Rösch den Begriff Generationsspektrum (gb).
Ein Beispiel: Hans Liebmann (ca. 1495 – 1570) tritt als siebenfacher Mehrfachahn sechs Mal in der 14. Ahnengeneration und einmal in der 15. Ahnengeneration des Probanden auf. In
der Notation von Rösch wird dies geschrieben als

Um dies auch statistisch zu erfassen, schlägt Rösch außerdem als Maß den Schwerpunktwert des biologischen Verwandtschaftsgrades (gbs) vor. Für Hans Liebmann ist gbs = 14,14. Dieser Wert verdeutlicht, dass Hans Liebmann deutlich „näher“ an der 14. Ahnengeneration (in der er sechs Mal auftritt) als an der 15. Ahnengeneration ist (in der er nur einmal auftritt).
„Erbwirksamkeit“ eines Mehrfachahnen
Genealogisch interessant ist auch die sog. Erbwirksamkeit eines Mehrfachahnen. Zunächst sind die Ahnen eines Probanden, die ihm näher liegen, biologisch von weitaus größerer Bedeutung als weit entfernte Ahnen, da sich der Gen-Anteil eines Probanden von einem Ahnen ja mit jeder weiteren Generation halbiert. Wie der deutsche Mathematiker und Autor mehrerer Schullehrbücher Hermann Athen (1911 – 1980) mathematisch zeigen konnte, ist die „Autosomen-Verwandtschaft“ nach „der 6. Generation eher unwahrscheinlich als wahrscheinlich“ (Athen, 1982, S. 424), da durchschnittlich nur noch 1/64 der Gene des Ahnen den Probanden erreichen. Es liegt aber auf der Hand, dass die biologische Bedeutung eines Ahnen für den Probanden umso größer ist, je häufiger er in der AT des Probanden auftaucht, da „seine Gene“ dann über viele (nämlich z > 1) verschiedene Nachfahrenlinien ihren Weg zum Probanden gefunden haben. Wie lässt sich also die (wahrscheinliche) Erbwirksamkeit eines Mehrfachahnen in einer bestimmten Generation mit der Erbwirksamkeit eines einfachen Ahns in einer bestimmten Generation vergleichen?
In Röschs Quantitativer Genealogie gibt auf diese Frage der sog. summarische biologische Verwandtschaftsgrad (abgekürzt mit g’b) eine Antwort. Er lässt sich über eine Logarithmus-Formel, einfacher jedoch mit der Software AhnenImplex berechnen. Ein Beispiel soll das verdeutlichen.
Der bereits oben erwähnte Hans Liebmann (ca. 1495 – 1570) tritt als siebenfacher Mehrfachahn sechs Mal in der 14. Ahnengeneration und einmal in der 15. Ahnengeneration des
Probanden auf. Mittels AhnenImplex lässt sich für ihn ein g’b-Wert von 11,29 (also abgerundet: 11) berechnen. Inhaltlich besagt dies, dass Hans Liebmann (der überwiegend nur in
der 14. Ahnengeneration auftritt) die gleiche Erbwirksamkeit wie ein einfacher Ahn in der 11. Ahnengeneration hat. Es besteht daher also eine größere Wahrscheinlichkeit dafür, dass evtl. ein
kleiner Teil seines Genoms auch im Genom des Probanden auftritt.
X-chromosomaler Erbgang
In der derzeit populären (und teilweise heftig umstrittenen) DNA-Genealogie können je nach Fragestellung unterschiedliche Arten von DNA untersucht werden: Die „normale“ autosomale DNA (atDNA), die DNA des Y-Chromosoms (yDNA), die nur über die rein väterliche Linie weitergegeben wird, die DNA des X-Chromosoms (xDNA) mit der Besonderheit, dass Männer ihr einziges X-Chromosom nur an ihre Töchter weitergeben können, nicht aber an ihre Söhne (wodurch unterschiedliche Häufigkeiten X-chromosomaler Ahnen in den Generationen entstehen), sowie die mitochondriale DNA (mtDNA), die nur über die rein mütterliche Linie vererbt wird (Abb. 5).
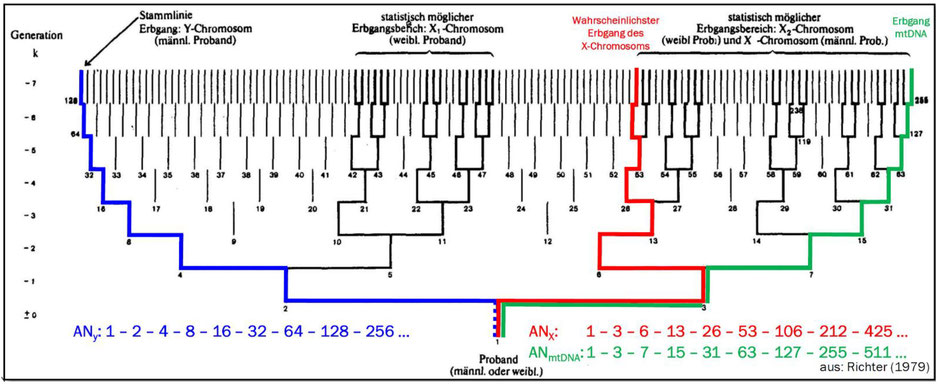
Abb. 5: Erbgang des Y-Chromosoms (blau) und der mitochondrialen DNA (grün) sowie die möglichen Erbgänge des X-Chromosoms (schwarz und rot) in einer AT (modif. nach Richter, 1979).
Siegfried Rösch hat 1955 die oben beschriebenen Berechnungsmethoden ausschließlich für den autosomalen Erbgang entwickelt. 1979 wurden diese durch Arndt Richter auf die X-chromosomale Verwandtschaft erweitert. Bemerkenswert ist hierbei, dass das X-Chromosom nur über wenige, spezifische Ahnenlinien an die Probandin bzw. den Probanden weitergegeben werden kann, da das X-Chromosom des Vaters ja nie an einen Sohn weitergegeben wird (Abb. 5).
Es macht allerdings einen Unterschied, ob der Proband ein Mann oder eine Frau ist: Bei Männern (die ihr X-Chromosom nur von der Mutter erhalten) kommen in der Ahnengeneration k = -6
dreizehn Ahnenlinien in Frage, die alle auf der mütterlichen Seite der Ahnentafel liegen, bei Frauen (die ihre beiden X-Chromosomen ja von Vater und Mutter erhalten) sind es 21
Ahnenlinien, da die acht Ahnenlinien der väterlichen Seite hinzukommen.

Abb. 6: Mögliche Erbgänge des X-Chromosoms in der AT Gerhard Kreile (AN = 1).
Mit Hilfe der Software AhnenImplex ist es möglich, eine „X-chromosomale Ahnentafel“ zu erstellen und die X-chromosomale Verwandtschaft näher zu analysieren (Abb. 6).
Hinweise zu "AhnenImplex"
- Fragen zum Programm bitte NICHT an den Programmentwickler oder den Roland zu Dortmund, sondern ausschließlich an Weert Meyer, den Autor des Handbuchs ([email protected])!
- Programm und Handbuch können hier kostenlos heruntergeladen werden.
- Da es sich bei AhnenImplex um eine ausführbare exe-Datei handelt, warnt Windows vor dem Download. Der Download kann jedoch bedenkenlos durchgeführt werden, das Programm wurde vor dem Upload auf Viren überprüft.
- Haftungsausschluss: Trotzdem erfolgen Download und Verwendung des Programms auf eigenes Risiko, für eventuelle Schäden wird keine Haftung übernommen.
- Das Programm darf ausschließlich zu privaten und nicht zu kommerziellen Zwecken verwendet werden.
(Dieser Blog-Beitrag kann mit Fußnoten und Literaturverzeichnis hier als pdf-Dokument herunergeladen werden.)